- Basic
- Publication (PDF)
- Software
- Example
- Imprint
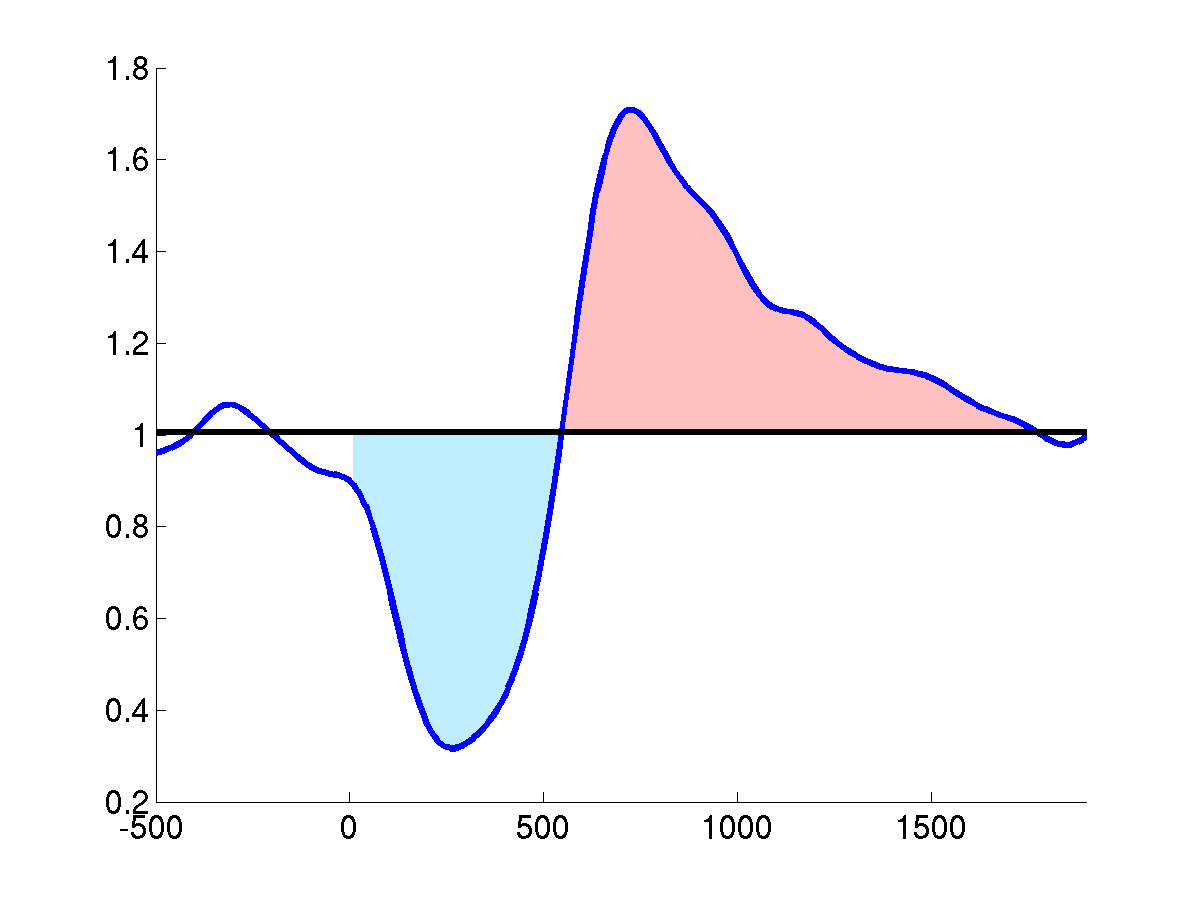
Somatosensory induced perturbation of mu-rhythm: after the
event-onset at t=0, the mu-rhythm desynchronizes, visible from the
decrease in averaged band power. This period of ERD (blue dyed area)
is followed by an increase in signal power, overshooting the pre-event
baseline level and indicating an ERS (red dyed area) and thus
yielding in the standard biphasic ERD-ERS response.
Mu-rhythm ERD: The human perirolandic sensorimotor cortices show rhythmic
macroscopic EEG/MEG oscillations with spectral peak energies around
10Hz (localized predominantly over the postcentral somatosensory
cortex) and 20Hz (over the precentral motor cortex). These
so-called mu-rhythms exhibit fast inherent fluctuations as they are
limited to brief periods (spindles) of 0.5-2sec duration, which
appear to occur in the absence of overtly processing sensory
information or motor commands. ERD/S of the mu-rhythm have been
reported for different physiological manipulations, e.g., by motor
activity, both actual and imagined, as well as by somatosensory
stimulation. In this context standard trial averages of
mu-rhythm power typically reveal a sequence of attenuation
followed by a rebound which often overshoots the pre-event baseline
level.
The study: In the following we give an example, how the framework of state conditional generalized ERD can be used to scrutinize explanatory factors and their impact on inter-trial variability of ERD responses. In particular we outline an investigation of the possible influence of local mu-rhythm activity prior to the event-onset on the magnitude and latency of induced mu-rhythm ERD. Additionally, we investigate the impact of occipital alpha activity. To this end, we present EEG data of a single subject from a somatosensory stimulation experiment, where mu-rhythm ERD was induced by electrical stimulations of median nerve at the right wrist.
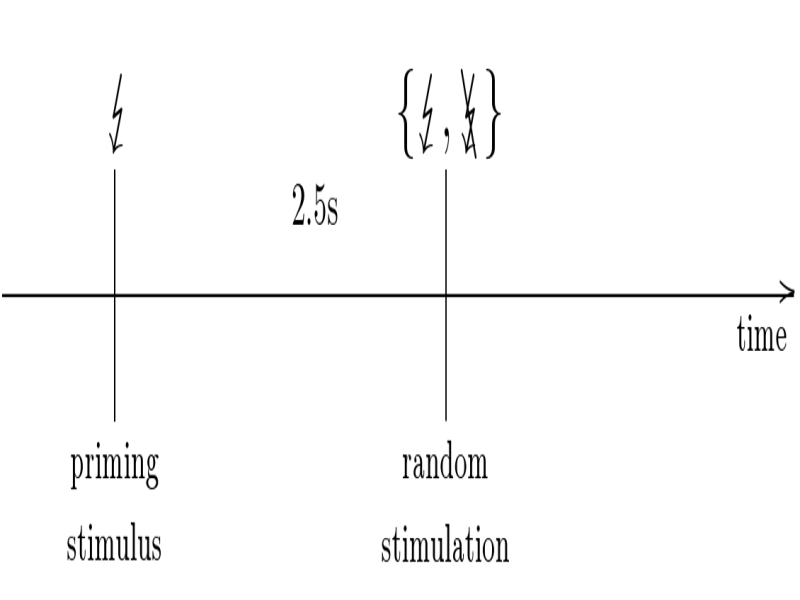
Single
trial layout: measures the relative deviation of the
event-related dynamics (solid) from a dynamic baseline (dashed). The
dynamic baseline can be estimated from catch trials, i.e., as averaged
power across trials without processing the event under study.
Experimental design: In order to
acquire an appropriate reference dynamics from catch trials, we used a
randomized stimulation paradigm, where a stimulus was delivered with
probability of 0,5, while with the same probability the stimulus was
refrained. The latter served to obtain catch trials of the natural
relaxation dynamics. Moreover, to mimic the condition of a repetitive
stimulation condition, each trial consists of a pair of stimuli. Here,
the first stimulus served as a priming stimulus and was always
delivered, while the second stimulus was delivered according the
random stimulation scheme. For the analysis of ERD inter-trial
variability only the responses to the second stimuli were considered.
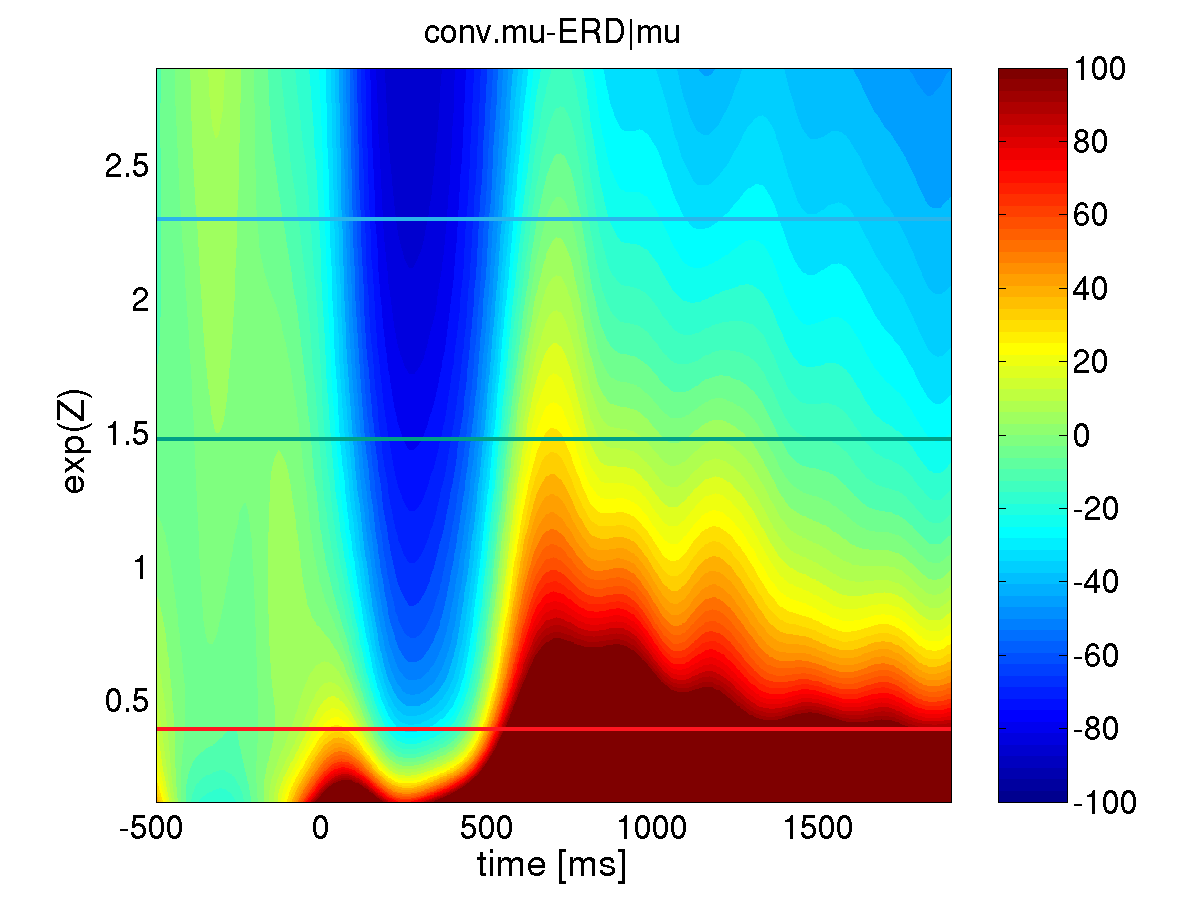
Conventional state conditional ERD: The estimated conditional ERD is depicted using a color coded scheme, where blue dyed areas correspond to periods of ERD while red dyed areas indicate ERS. The vertical axis refers to the pre-stimulus activity of the local mu-rhythm. Depending on the level of activity (and therefore depending on the baseline level) the conventional ERD analysis yields a rather questionable, variable result, suggesting only an ERD response at high levels of activity and only an ERS at low levels activity.
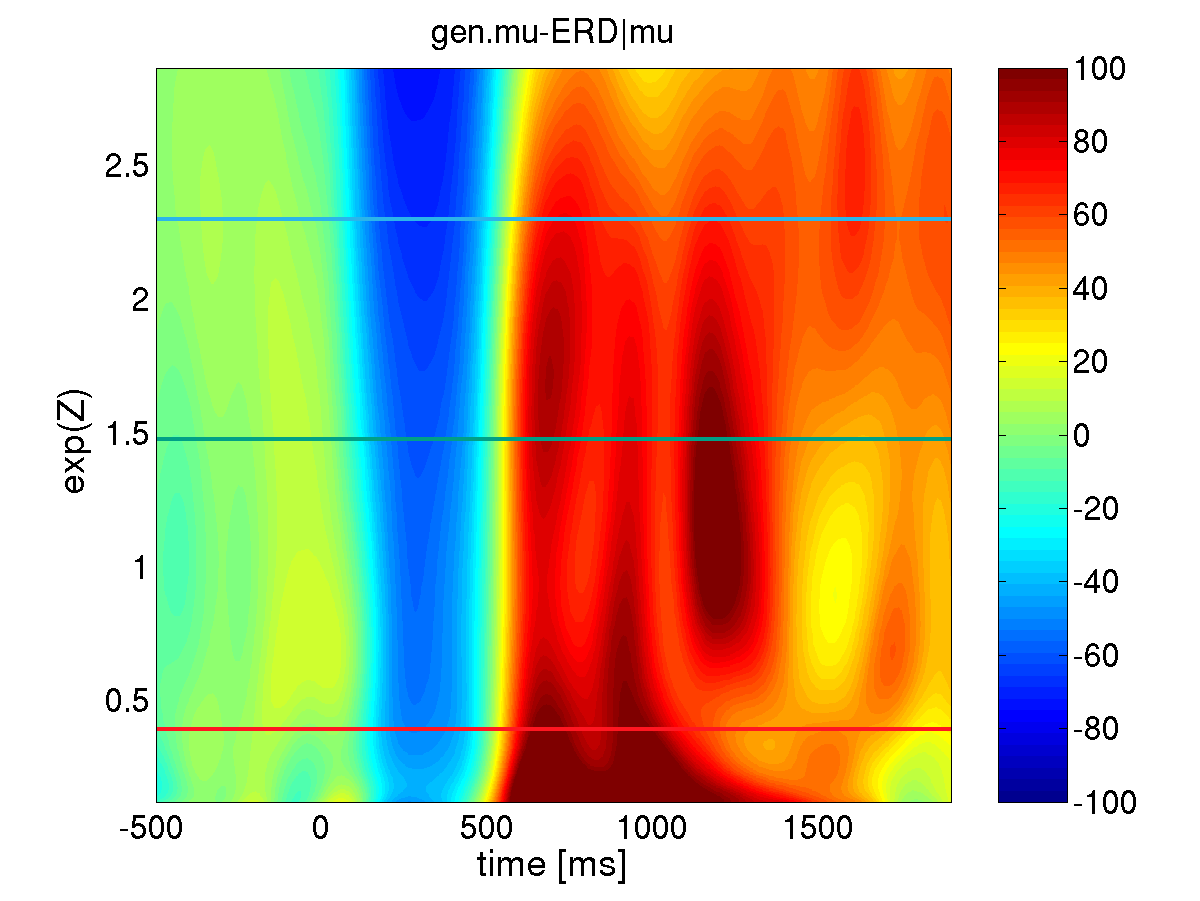
Generalized state conditional ERD: The estimated conditional ERD is depicted using a color coded scheme, where blue dyed areas correspond to periods of ERD while red dyed areas indicate ERS. The vertical axis refers to the pre-stimulus activity of the local mu-rhythm. The generalized state conditional ERD analysis reveals that the human brain reacts in a highly systematic biphasic ERD/S sequence independent of the preceding state with either low or high amplitudes of ongoing mu-oscillations. Comparing generalized and conventional ERD: As we want to scrutinize the impact of the local mu-rhythm and alpha activity immediately preceding the event, we defined the explanatory variable Z as, the logarithm of the averaged mu-rhythm (alpha) power across the interval [-300;-100]ms prior to the second stimulus. Applying both, the conventional and the generalized state conditional ERD measures with respect to the impact of local mu-activity, it turns out that only the generalized method is able to reliably disclose the standard biphasic ERD-ERS complex (lower panel) and therefore allows for further analysis of the state conditional functional relationship between mu-rhythm ERD and the local pre-stimulus mu-activity. Moreover, the conventional framework gives rise to the observation of solely ERD (upper area in the upper panel) or solely ERS (lower area in the upper panel).
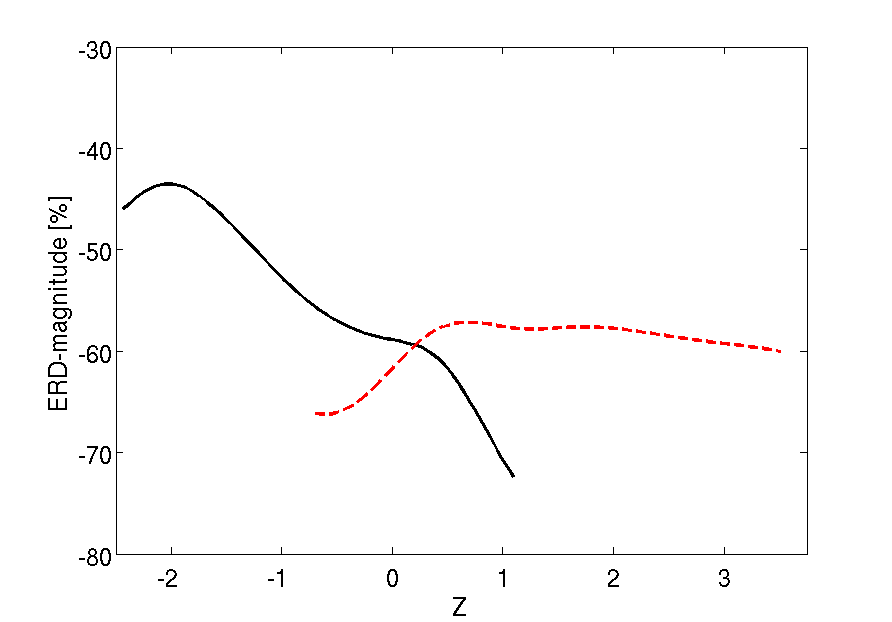
Dependency of ERD magnitude on the two investigated explanatory factors, i.e., the local mu-activity (solid black) and the occipital alpha activity (red dashed). Analysis of a significant monotonic relationship reveals, the higher the pre-stimulus mu-activity the stronger the mu-ERD.
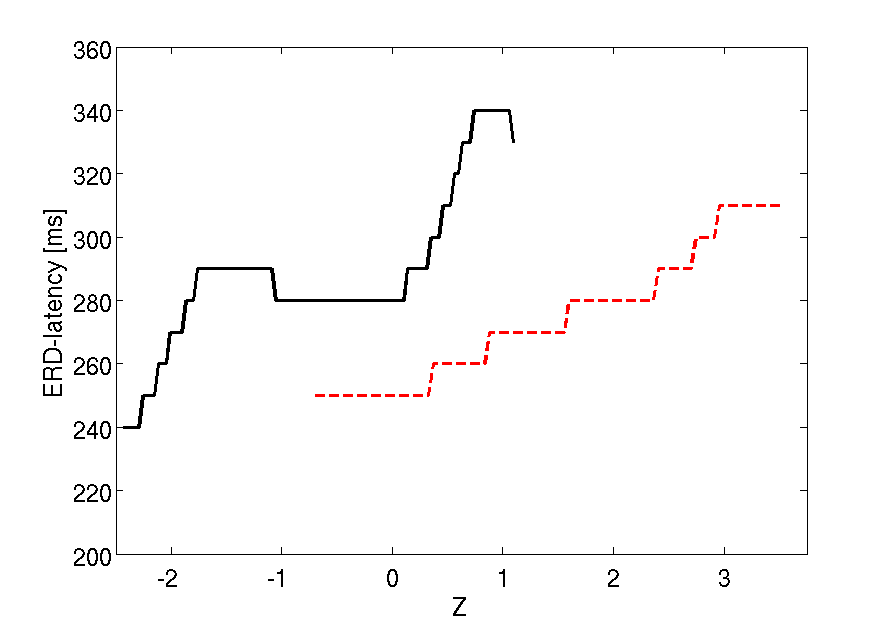
Dependency of ERD latency on the two investigated explanatory factors, i.e., the local mu-activity (solid black) and the occipital alpha activity (red dashed). A significant monotonic relationship was found with respect to the impact of alpha activity. Particularly, the higher the pre-stimulus occipital alpha activity the later the mu-ERD appears. Explanatory factors of ERD inter-trial variability: Restricting the analysis of state conditional ERD to the generalized measured only, we further investigated the functional relationship of the magnitude of local mu and occipital alpha pre-stimulus activity on the one hand and the magnitude and latency of deepest ERD on the other hand. Here, the application of the novel measure provided evidence for the existence of a sensible physiological dynamics related to the interaction between ongoing activity and stimulus induced responses. In particular, we found a significant monotonic negative relationship between the magnitude of ERD and the local mu-activity, while the occipital alpha revealed no effect on the magnitude of mu ERD. Physiologically, these results show that the ERD is stronger in case of immediately preceding higher pericentral mu-activity and can therefore be considered as a strictly local phenomenon. On the other hand, the latency of the ERD response showed a positive monotonic relationship with occipital alpha activity; Thus, a lower occipital alpha, possibly indicative of system-wide increase of arousal, is reflected by a faster ERD at pericentral cortices.
Copyright © 2009 S. Lemm, K.-R. Müller, and G. Curio.
The original work was published under the Open Access License, which allows anyone to download, reuse, reprint, modify, distribute, and/or copy articles in PLoS journals, so long as the original authors and source are cited. If the item you plan to reuse is not part of a published article (e.g. a featured issue image), then please indicate the originator of the work, and the volume, issue and date of the journal in which the item appeared.For any reuse or redistribution of a work, you must also make clear the license terms under which the work was published.